

Après avoir rappelé le contenu du terme « biodiversité », cet article présente les connaissances relatives aux variations de la biodiversité, en France, entre le dernier maximum glaciaire et le maximum holocène. Il fournit ensuite quelques exemples d'impacts du réchauffement climatique actuel sur les écosystèmes et termine par une comparaison entre les réchauffements passé et actuel, permettant une projection vers le futur.
Introduction
La biodiversité
La biodiversité peut être définie comme l’ensemble de la diversité du vivant, à savoir :
- la variabilité génétique au sein de chaque espèce,
- la diversité des espèces et
- la variété des écosystèmes.
Robert Barbault préfère parler de « tissu vivant de la planète » pour mieux prendre conscience des interactions.
Le réchauffement climatique
Le réchauffement actuel de la planète est un fait : entre 1900 et 2007, la température moyenne du globe a augmenté de 1 °C (15 °C de température moyenne pour 2006) et le niveau moyen des océans est monté de 15 cm.
La Terre a connu de nombreuses variations climatiques avec des fluctuations de température de ± 15 °C par rapport à l’actuel et des changements du niveau marin d’au moins ± 200 m au cours des 650 derniers millions d’années.
À une échelle plus « humaine », le dernier maximum glaciaire date de 18 ka (1 ka = 1000 ans), période à laquelle la température moyenne de la Terre était alors de 4 à 5 degrés inférieure à l’actuelle. En seulement 5 000 ans, le principal de la déglaciation était fait. Durant cette tendance générale, après la première période de forte déglaciation, plusieurs petits épisodes chauds ou froids ont eu lieu.
Le passé : dernière déglaciation et biodiversité
Le dernier maximum glaciaire
Le dernier maximum glaciaire (DMG) a 18 ka. Un réchauffement climatique général a ensuite entraîné une déglaciation continue jusqu’à aujourd’hui, avec un maximum de réchauffement il y a environ 8 ka (maximum holocène, ou MH). La température moyenne au MH est de l’ordre de 2 °C supérieure à l’actuelle.
Ainsi, la différence de température moyenne entre ces deux évènements est-elle de l’ordre de 6 à 7 °C. Mais, une différence de 7 °C ne signifie pas que cette différence est la même partout.
Les variations de température sont beaucoup plus faibles à l’équateur qu’aux moyennes, et encore plus, qu’aux hautes latitudes. Ainsi, dans le Nord-Est de la France, peut-on estimer le réchauffement de 16 à 22 °C entre le DGM et le MH.
Comparaison de la faune et de la flore en France entre – 18 ka et – 8 ka
Des « Cartes des environnements du monde pendant les deux derniers extrêmes climatiques (CLIMEX) » ont été publiées conjointement par l’ ANDRA (Agence nationale pour la gestion des déchets radioactifs) et la CCGM (Commission de la carte géologique du monde).
L’ANDRA propose aussi l’équivalent pour le bassin méditerranéen.
Nous nous intéresserons ici aux cartes de la France lors des deux derniers extrêmes climatiques (cartes ANDRA / CNF-INQUA / IGN), dont le CNRS publie aussi des commentaires sur les grands mammifères, entre le dernier épisode glaciaire et l’Holocène.

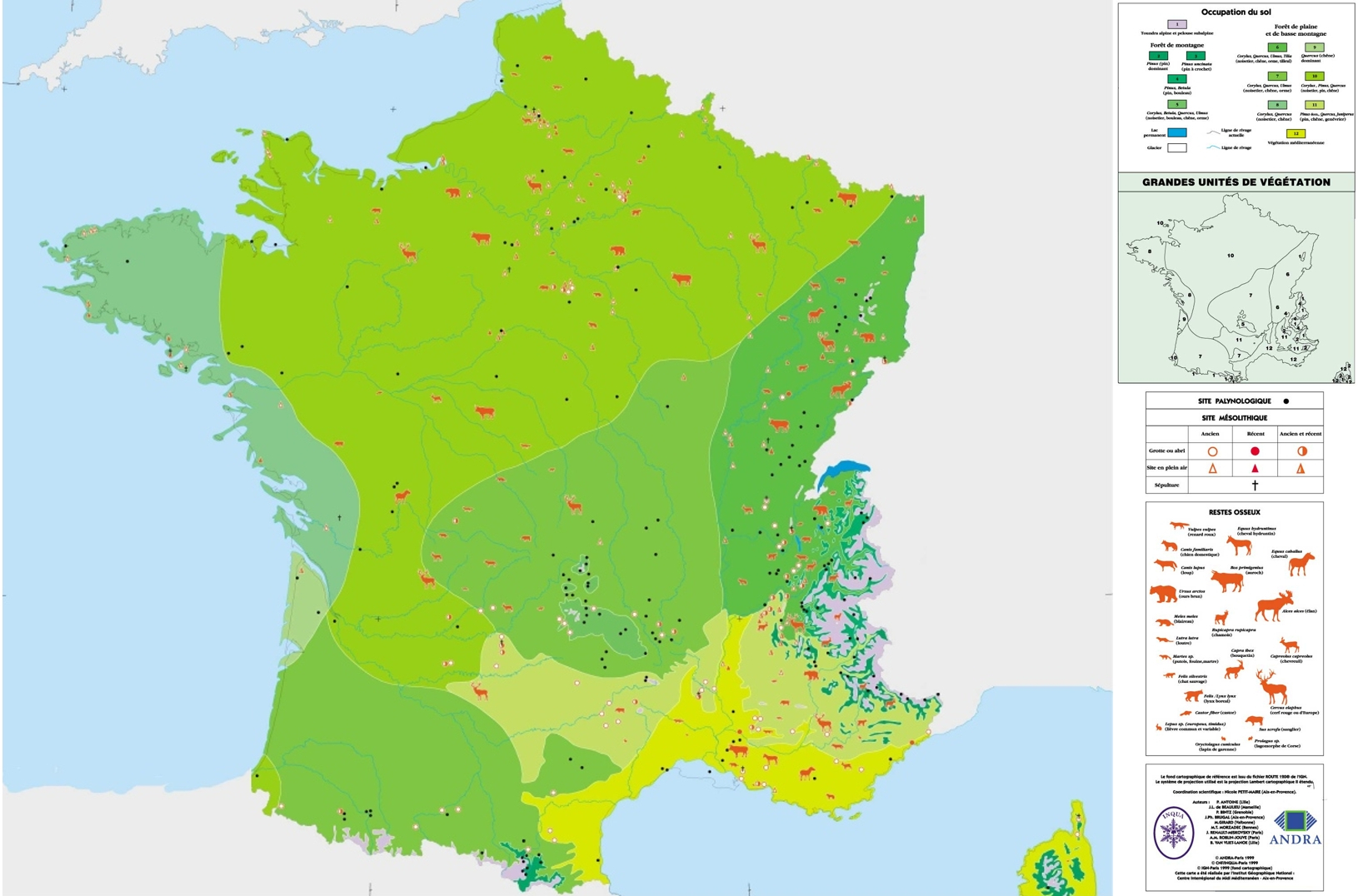
Entre le DMG et le MH, le niveau de la mer est remonté de -120 m à -25 m par rapport à l’actuel. En effet, des calottes glaciaires ayant pu persister en Amérique du Nord jusqu’à -5 ou -6 ka, le niveau marin au MH est plus bas qu’à l’actuel, pourtant plus froid.
Ceci a pour conséquence d’isoler la Grande-Bretagne du reste du continent, mais, surtout, les sols dégèlent : glaciers et pergélisols disparaissent des plaines.
Certains végétaux ou associations végétales disparaissent du territoire (comme la steppe), d’autres voient leur extension limitée aux zones d’altitude (comme la toundra alpine ou le pin). Les espèces « disparues » perdurent cependant aujourd’hui plus au Nord.
Concernant les animaux, certains disparaissent de France mais perdurent dans les régions plus septentrionales, voire boréales actuelles : le phoque moine, le renne, le renard polaire, par exemple. D’autres animaux disparaissent entre ces deux périodes : le mammouth, le lion des cavernes, la hyène des cavernes. Certains animaux ont ainsi migré en même temps que se déplaçaient les écosystèmes favorables. Les causes de la disparition du mammouth il y a environ 12 ka, bien que discutées, semblent elles aussi liées aux changements climatiques ; la prédation par l’Homme étant à l’époque vraisemblablement beaucoup trop limitée pour causer l’extinction de l’espèce.
On peut déjà remarquer que l’échelle à laquelle on se place est importante. La disparition locale d’une espèce ne signifie pas nécessairement baisse de la biodiversité, si celle-ci perdure par ailleurs suite à une migration ou du fait de l’existence d’une occupation spatiale initialement très vaste. Cependant, certaines espèces ont bel et bien disparu lors de cette phase de réchauffement climatique.
L’adaptation des écosystèmes aux variations climatiques dépend, entre autres, de la capacité de mobilité de ses constituants. Pour les animaux, la mobilité est liée à la mobilité des individus et est alors limitée par les barrières naturelles (fleuves, montagnes). Chez les végétaux, bien que les individus soient immobiles, la mobilité des espèces est assurée lors de la reproduction (nombreuses spores ou graines « voyageuses »).
Ainsi, le réchauffement moyen entre le DMG et le MH (6 à 7 °C à l’échelle globale, et environ 20 °C à l’échelle de la France, en 10 ka) a été suffisamment important pour modifier en profondeur la répartition des écosystèmes, mais certainement suffisamment « lent » pour maintenir la biodiversité. Seules les espèces les plus fragiles ont pâti de ce réchauffement climatique général.
L’actuel : quelques exemples de l’influence du réchauffement climatique actuel sur les écosystèmes
La température est un paramètre abiotique fondamental qui règle de nombreux processus du vivant. Le climat, qui intègre la température ainsi que de nombreuses autres variables abiotiques, elles aussi, influence de manière très importante la vie des organismes et les relations que ces organismes entretiennent dans le cadre des écosystèmes.
Connaissant ce forçage climatique capital, on comprend qu’une modification du climat telle que nous la connaissons actuellement, c’est-à-dire une augmentation à la fois des températures moyennes, mais aussi de la variabilité des températures autour de la moyenne, soit susceptible d’avoir de lourdes conséquences sur les individus, les espèces et les communautés.
La littérature abordant le sujet se fait de plus en plus abondante et nous allons ici présenter quelques exemples des influences climatiques sur les êtres vivants et les écosystèmes.
Modifications des aires de répartition
Parmi les premiers symptômes du réchauffement climatique reconnus en tant que tels, on trouve tout d’abord les modifications d’aires de répartition. En effet de nombreux organismes, notamment ectothermes, ont des limites septentrionales et méridionales de répartition géographique fixées par des isothermes.
Les papillons, par exemple, sont connus comme étant généralement très sensibles au paramètre température. Ils ont donné lieu à une étude princeps sur les déplacements des aires de distribution. Parmesan et al. (1999) [1] ont ainsi démontré que, sur 35 espèces de papillons sédentaires de Grande-Bretagne, 63 % avaient vu leurs aires de répartition progresser vers le Nord, de 35 à 240 km. Un exemple de ces déplacements d’aire de répartition est présenté sur la figure suivante pour une espèce de Nymphalinés, le Tircis (Pararge aegeria).
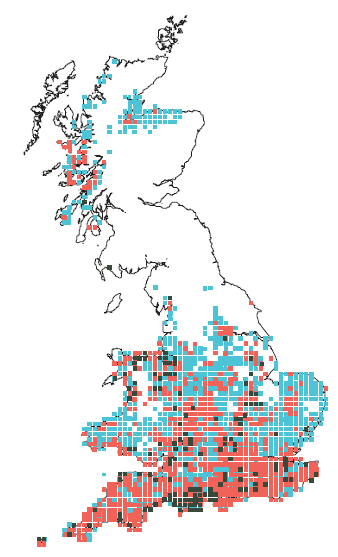
Les cellules indiquent la présence d'au moins une population de tircis relevée lors d'un suivi national entre 1915 et 1939 (vert foncé), entre 1940 et 1969 (rouge) et entre 1970 et 1977 (bleu).
On constate une extension de l'aire de répartition vers le nord.
Ce type d’étude a concerné tous types de taxons et il a été montré, par exemple, que la croissance végétale avait augmenté aux hautes latitudes (Myneni et al, 1997) [2]. Il faut remarquer que les déplacements d’aires mis en évidence représentent des dynamiques de colonisations/extinctions en limite de distribution ou des déplacements « réels » d’individus par migration.
Or toutes les espèces ne sont pas capables de suivre les modifications de leur environnement dues au changement climatique par la colonisation de nouveaux espaces, parce que d’autres paramètres règlent leur répartition. Par conséquent, cela se traduit par des déclins d’espèces, pour celles qui ne peuvent pas répondre, et par des progressions, pour celles qui y répondent le mieux.
Un autre exemple de modifications récentes des aires de répartition de certaines espèces ou de modifications de la biodiversité spécifique en un lieu donné (dont tout laisse à penser qu’elles sont en relation avec la modification du climat) est fourni avec les diatomées. Une étude de Coste et Ector (2000) [3] a ainsi montré que, depuis 20 à 30 ans, il y a eu apparition et/ou prolifération, dans les cours d’eau français, de plusieurs espèces de diatomées, notamment d’origines tropicales ou subtropicales, considérées comme de bons indicateurs de l’augmentation de la température de l’eau de surface.
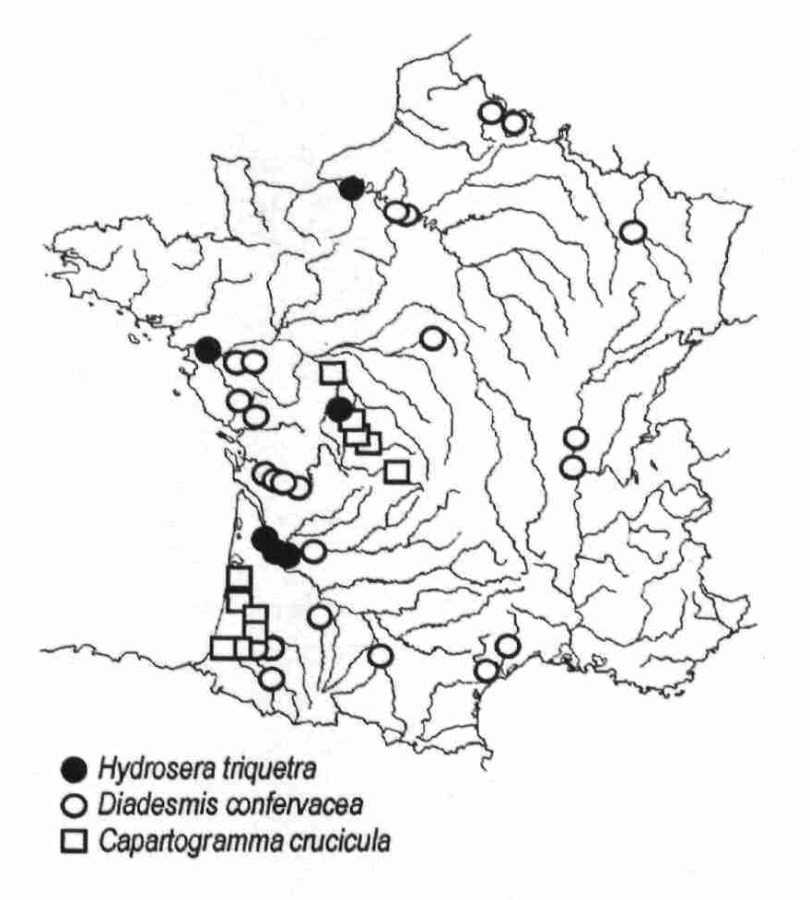
L’arrivée de nouvelles espèces de diatomées en France peut avoir différentes origines qui ne sont pas nouvelles : l’eau et la pluie, les insectes aquatiques, les oiseaux migrateurs, la navigation et les activités humaines, les espèces végétales ou animales introduites. En revanche, pour que les nouvelles espèces puissent s’acclimater, il faut qu’elles y trouvent les conditions nécessaires à leur survie, c’est ce qui en fait de bons indicateurs d’éventuelles modifications de l’environnement. Pour citer un exemple précis, on parlera de Diadesmis confervacea (figure ci-dessous), diatomée thermophyle considérée comme étant d’origine tropicale, et qui est à présent largement distribuée de par le monde, y compris dans les régions tempérées. Décrite en Europe depuis plus d’un siècle dans des lieux particuliers tels des serres de jardins botaniques ou des sources thermales chaudes, elle est décrite pour la première fois en France, durant les étés 1972 à 74, dans les rejets d’eau chaude de la centrale thermique EDF de Porcheville, donc en Île-de-France, puis en 1976 dans la Loire, en aval de la centrale nucléaire de St-Laurent-des-Eaux près d’Orléans. On voit donc clairement l’association entre cette diatomée et la température de l’eau. Depuis les années 90, cette espèce est régulièrement décrite dans de nombreux cours d’eau, le plus souvent situés au sud de la Loire, non nécessairement associée avec une source locale d’eau chaude (Adour, canal de la Robine, Charente, Dordogne, Isle, Canal du Jard, Indre, Loire [site EDF de Saint-Laurent-des-Eaux], Moine, Vendée).
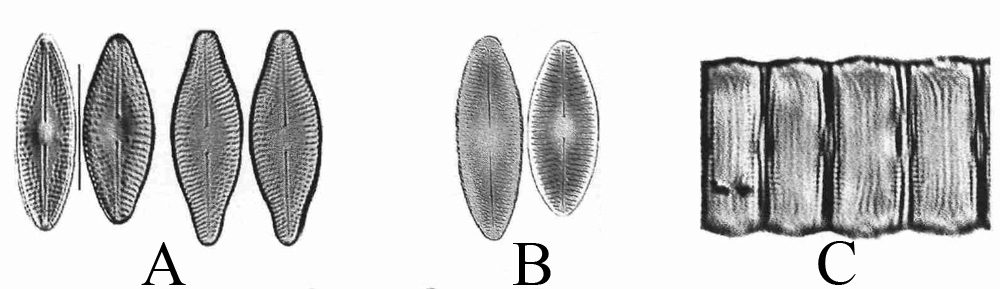
Les photos A sont des vues valvaires prises en microscopie optique (MO), les photos B ont été prises en microscopie électronique à transmission (MET) et les photos C sont des vues connectives prises en MO.
D’autres espèces tropicales thermophiles telles Hydrosera triquetra et Capartogramma crucicula (voir figures ci-dessous ) se sont également implantées en France assez récemment.
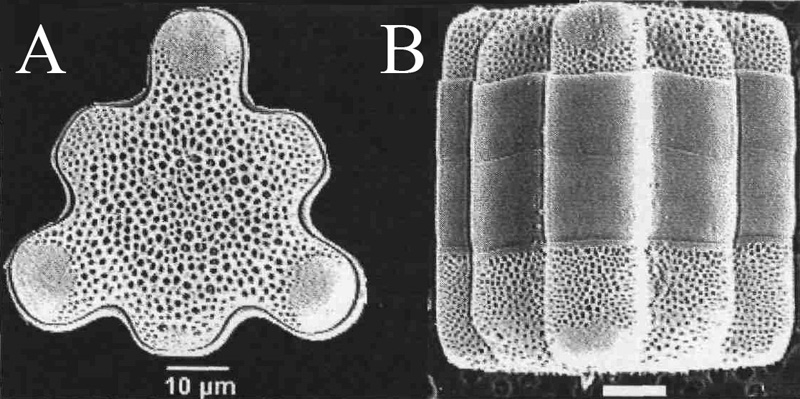
Vues valvaires et connective prises en microscopie électronique à balayage (MEB).
Les photos A ont été prises en MO, la photo B en MET.
La figure ci-dessous montre la distribution observée en France pour ces trois espèces.
Modification des cycles de vie
En second lieu, le réchauffement climatique actuel entraîne des modifications dans la disposition temporelle des étapes du cycle de vie des organismes.
Un exemple connu est la conséquence d’un printemps chaud sur la précocité de floraison des arbres. Menzel, Estrella et al. (2001) [4] ont observé une avancée dans le débourrement des bourgeons et la floraison de 1,4 à 3,1 jours, par décennie, sur une cinquantaine d’années.
En moyenne, on constate depuis quelques décennies que la phénologie des êtres vivants, c’est-à-dire le positionnement dans l’année et la durée des phases du cycle de vie, répond au changement climatique.
Ainsi, dans leur revue sur les conséquences générales du réchauffement climatique sur les écosystèmes, Parmesan et al. (2003) [5] présentent l’évolution de la date moyenne de phénophases de différents organismes (la phénophase désigne les synusies, ou développement de certains êtres vivants qui se met en place en relation avec une période particulière de l’année). On constate un avancement graduel dans le temps de ces phénophases (figure ci-dessous).
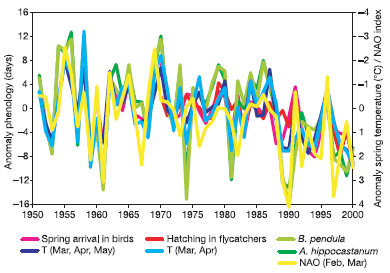
Anomalies temporelles = variations par rapport à la moyenne.
Les phénophases : retour migratoire printanier des oiseaux (rose), date de ponte du gobemouche noir Ficedula hypoleuca (rouge), débourrement des bourgeons chez le marronnier commun Aesculus hippocastanum (vert foncé) et le bouleau verruqueux Betula pendula (vert clair).
Les variables climatiques : températures moyennes (mars, avril, mai) en bleu foncé, températures moyennes (mars, avril) en bleu clair et NAO moyen de février et mars en jaune.
Dans le cas des modifications phénologiques (étude de l’apparition d’événements périodiques déterminée par les variations saisonnières du climat) du cycle de vie des organismes, on trouve aussi des espèces capables de répondre aux modifications en avançant leur phénophase et d’autres qui en sont moins capables.
Ainsi, les espèces qui, faute de pouvoir avancer leur phénologie, se retrouvent en décalage vis-à-vis de leur ressource, connaissent un déclin. C’est par exemple le cas chez les oiseaux migrateurs qui parviennent difficilement à ajuster leur date de retour migratoire, laquelle est contrainte par de nombreux paramètres indépendants de la température sur le lieu de reproduction.
Chez les oiseaux le cas du gobemouche noir (Ficedula hypoleuca) a été bien documenté.
Both et al. (2006) [6] ont montré sur différentes populations hollandaises de cet oiseau migrateur trans-saharien que la tendance démographique à long terme était corrélée à la date du pic d’abondance de chenilles, dont dépend cet oiseau insectivore, et au coefficient de corrélation entre la date de ponte moyenne de la population et la température printanière (figure ci-dessous).

On constate que les populations en déclin sont celles qui subissent un pic très précoce de leurs ressources et qui n’arrivent pas à pondre plus tôt (coefficient de régression plus proche de 0).
Enfin, nous avons constaté que le sort démographique des espèces face au changement climatique ne dépendait pas uniquement de l’espèce elle-même mais plus largement de toutes les espèces ayant des liens communautaires avec elle. En d’autre termes, pour bien mesurer les conséquences du réchauffement climatique sur la biodiversité, il faut tenir compte des réponses relatives des espèces et des liens de prédation, de compétition, de facilitation, qui les unissent au sein des écosystèmes en des réseaux trophiques complexes.
L’exemple du gobemouche est parlant : pour maintenir une démographie stable, il doit avancer sa date de ponte d’un temps équivalent à celui de l’avancée du pic de ressource (ici les chenilles dont il dépend), ce qui représente davantage qu’un simple avancement graduel sans lien avec la communauté.
L’écologie du réchauffement climatique se fait, se conçoit désormais, à l’échelle de la communauté d’espèces, mais il reste encore beaucoup à faire pour comprendre toutes les répercussions des modifications, à l’échelle d’une espèce, sur l’ensemble de la communauté.
Conséquences du changement climatique visibles en France
Pour terminer, considérons les conséquences telles qu’elles sont appréhendées en France.
Les oiseaux sont considérés comme de bons indicateurs des changements globaux en général et des changements climatiques en particulier. En effet, ils sont situés à un niveau assez haut dans les chaînes trophiques et intègrent par conséquent de nombreux paramètres de la modification de leurs environnements.
Il existe en France un observatoire national de la biodiversité s’intitulant « Vigie Nature », dont le fer de lance est constitué par le programme de suivi temporel des oiseaux communs (STOC). Le Muséum National d’Histoire Naturelle, qui centralise ces données, contribue beaucoup à l’étude des conséquences des changements globaux et du réchauffement climatique, en particulier sur la biodiversité.
Ainsi, Jiguet et al. (2006) [7] ont, par exemple, étudié les conséquences de l’importante canicule de 2003 sur les taux de croissance des populations d’oiseaux, afin de mieux comprendre quelles espèces, dans un avenir plus chaud, seraient les plus affectées. Ils ont mis en évidence que les espèces qui avaient le plus souffert étaient celles dont les aires de distribution avaient les amplitudes thermiques les plus faibles (figure ci-dessous).
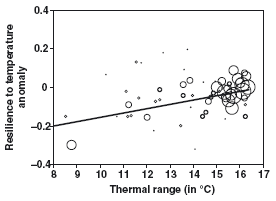
Relation entre la résilience à l’anomalie de température (= coefficient de régression entre la variation du taux de croissance d’une espèce entre 2003 et 2004 et l’anomalie de température estivale) et l’amplitude thermique de l’aire de répartition de l’espèce.
On constate que les espèces qui ont une amplitude thermique importante sur leur aire de distribution ont mieux résisté à la canicule de 2003.
Le futur : conséquences certaines… précision incertaine
Les prévisions du GIEC pour le réchauffement durant le siècle à venir sont de l’ordre de 1,8 à 4 °C, selon les scénarios envisagés pour le développement des activités humaines. Cela nous mène à un réchauffement minimal de près de 3 °C en 2 siècles (1900-2100).
Comparé aux 6 à 7 °C, à l’échelle globale, entre le DMG et le MH, soit en 10 000 ans, le réchauffement actuel (1,5 °C par siècle en moyenne) semble beaucoup plus rapide que pour l’intervalle DMG-MH (0,07 °C par siècle en moyenne).
Cependant, une moyenne sur 10 ka ne peut être directement comparée à une moyenne sur 2 siècles, car elle intègre des fluctuations « chaud-froid » qui peuvent masquer des périodes de réchauffement plus rapides. En effet, des variations de température importantes, de l’ordre de 16 °C en 150 ans, ont été estimées lors d’un épisode daté à -70 ka, lors de la dernière période glaciaire, mais au Groenland (A. Landais et al., 2004) [8], donc à de hautes latitudes plus sensibles aux variations globales et répondant ici à des phénomènes non-linéaires (non proportionnalité entre la variation de température moyenne de la Terre et la variation locale résultante). Ainsi, même si les variations de température moyenne de la Terre dans le passé ne sont pas aisées à obtenir sur de petites périodes, le réchauffement climatique en cours approche, voire dépassera, la vitesse de 2 °C par siècle, qui, selon certains spécialistes, est un seuil critique au-delà duquel de nombreuses espèces vivantes ne peuvent assurer leur survie en migrant.
Même avec des prévisions fiables sur les variations de température à l’échelle de la Terre, il reste très difficile de prévoir les impacts locaux avec autant de précision. Cependant, grâce à une meilleure compréhension des phénomènes météorologiques et climatiques actuels, les grandes lignes des changements climatiques régionaux peuvent être déterminées.
La biodiversité diminuera très certainement du fait de la vitesse du réchauffement actuel, mais plus une espèce a une aire de répartition vaste (surtout en latitude) et une population importante, plus ses chances de survie seront importantes.
En ce qui concerne l’espèce humaine, elle est aujourd’hui adaptée à presque tous les écosystèmes (sauf les plus extrêmes) et sa population est importante. Biologiquement, l’espèce a de très bonnes chances de survie, a priori, mais les difficultés, si elles ne sont pas biologiques, seront liées aux « sociétés humaines ». Ainsi, ce sont elles qui devront s’adapter et faire face (en plus de la crise énergétique en cours) au réchauffement climatique et à ses conséquences : immigration « climatique », conflits territoriaux, bouleversement des économies (ressources agricoles), montée du niveau marin, augmentation de l’intensité et de la fréquence des aléas climatiques…
Référence
- C. Parmesan, N. Ryrholm, C. Stefanescu, J.K. Hill, C.D. Thomas, H. Descimon, B. Huntley, L. Kaila, J. Kullberg, T. Tammaru, W.J. Tennent, J.A. Thomas, M. Warren. 1999. Poleward shifts in geographical ranges of butterfly species associated with regional warming. Nature 399, 579-583
- R.B. Myneni, C.D. Keeling, C.J. Tucker, G. Asrar, R.R. Nemani. 1997. Increased plant growth in the northern high latitudes from 1981 to 1991. Nature 386, 698-702
- M. Coste, L. Ector. 2000. Diatomées invasives exotiques ou rares en France : principales observations effectuées au cours des dernières décennies. Syst. Geogr. Pl. 70, 373-400
- A. Menzel, N. Estrella, P. Fabian. 2001. Spatial and temporal variability of the phenological seasons in Germany from 1951 to 1996 . Global Change Biology 7, 657-666
- C. Parmesan, G. Yohe. 2003. A globally coherent fingerprint of climate change impacts across natural systems. Nature 421, 37-42
- C. Both, S. Bouwhuis, C.M. Lessells, M. Visser. 2006. Climate change and population declines in a long-distance migratory bird. Nature 441, 81-83
- F. Jiguet, R. Julliard, C.D. Thomas, O. Dehorter, S.E. Newson, D. Couvet. 2006. Thermal range predicts bird population resilience to extreme high temperatures . Ecology Letters 9, 1321-1330
- A. Landais, N. Caillon, J. Severinghaus, J.M. Barnola, C. Goujon, J. Jouzel, V. Masson-Delmotte. 2004. Analyse isotopique de l’air piégé dans la glace pour quantifier les variations de température . C. R. Geoscience 336, 963–970




